

Post by thesporerex on Feb 17, 2014 23:00:02 GMT
Allosaurus fragilis

Scientific classification
Kingdom: Animalia
Phylum: Chordata
Class: Reptilia
Superorder: Dinosauria
Order: Saurischia
Suborder: Theropoda
Clade: Carnosauria
Family: Allosauridae
Subfamily: Allosaurinae
Genus: Allosaurus
Species: Allosaurus fragilis
Scientific classification
Kingdom: Animalia
Phylum: Chordata
Class: Reptilia
Superorder: Dinosauria
Order: Saurischia
Suborder: Theropoda
Clade: Carnosauria
Family: Allosauridae
Subfamily: Allosaurinae
Genus: Allosaurus
Species: Allosaurus fragilis
Fossil range: Late Jurassic Kimmeridgian to early Tithonian, 155-150 million years ago
Length: ~8.5-9.7 metres long(?12 metres) ~9+ metres on average.
Weight: ~1.5-?3 tons? (?5 tons)
Height:(Unpublished without kangaroo pose)~2-2.7 metres tall (based on comparisons)
Location: North america, Portugal and ?Tanzania
Skull length: 80cm-?1 metre?
Skull length: 80cm-?1 metre?
Name meaning: "different lizard"
Description:
Description:
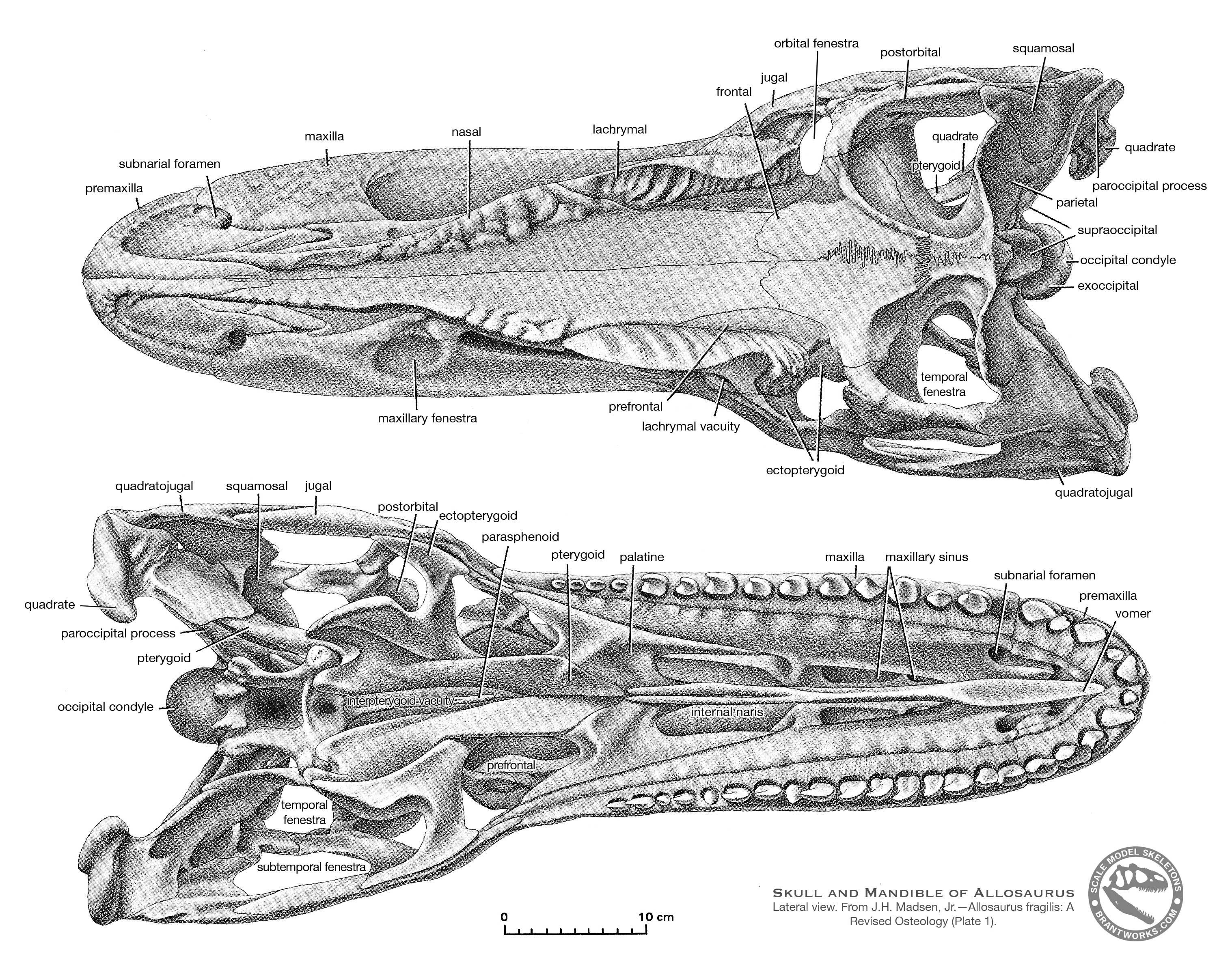
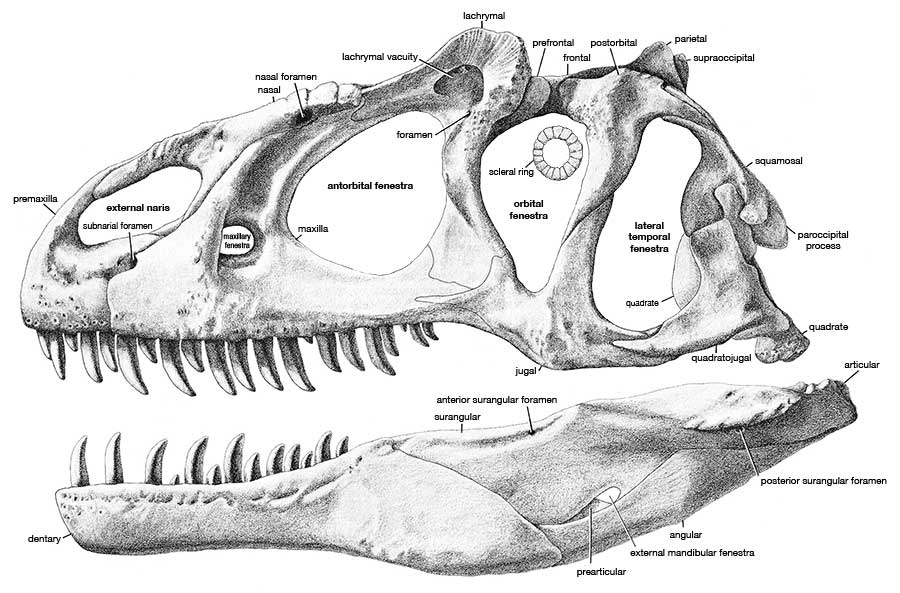
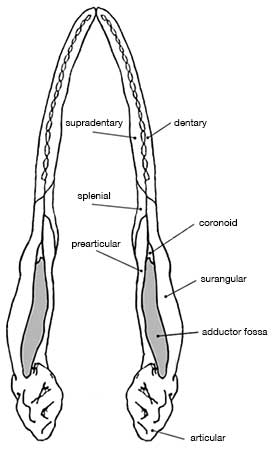
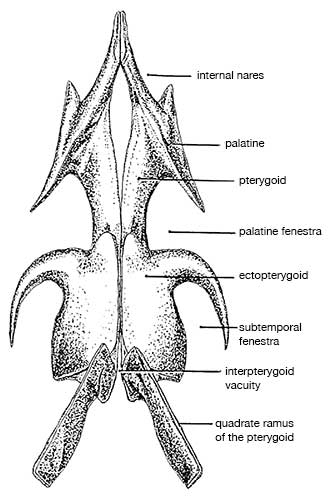
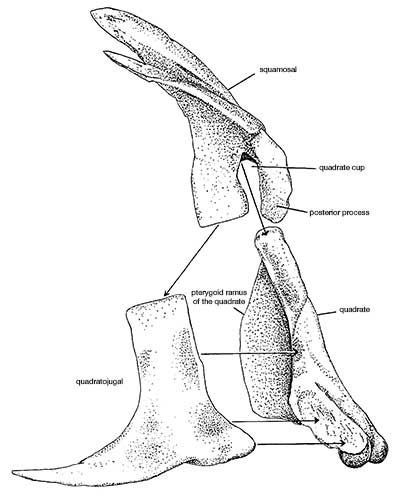
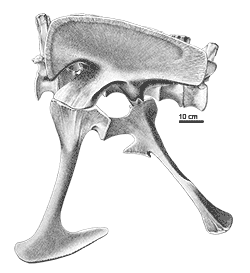
Allosaurus was a typical large theropod, having a massive skull on a short neck, a long tail and reduced forelimbs. Its most distinctive feature was a pair of blunt horns, just above and in front of the eyes. Although short in comparison to the hindlimbs, the forelimbs were massive and bore large, eagle-like claws. The skull showed evidence of being composed of separate modules, which could be moved in relation to one another, allowing large pieces of meat to be swallowed. The skeleton of Allosaurus, like other theropods, displayed bird-like features, such as a furcula (wishbone) and neck vertebrae hollowed by air sacs.
Allosaurus fragilis had an average length of 7-9 meters (~30 feet), with the largest definitive Allosaurus specimen (AMNH 680) measuring 9.7 m (32 ft). Several gigantic specimens have been attributed to Allosaurus, but may in fact belong to other genera. The closely related genus Saurophaganax (OMNH 1708) reached 10.9 m (36 ft) in length, and has sometimes been included in the genus Allosaurus as Allosaurus maximus, though recent studies lend support to the idea that it does belong in a seperate genus. Another specimen, once assigned to the genus Epanterias (AMNH 5767), may have measured 12.1 m in length. Epanterias may be a species of Allosaurus or Saurophaganax.
As with dinosaurs in general, weight estimates are debatable, and since 1980 have ranged between 1500 kilograms (3300 lb), 1000 to 4000 kilograms (2200 to 8800 lb), and 1010 kilograms (2230 lb) for modal adult weight (not maximum). John Foster, a specialist on the Morrison Formation, suggests that 1000 kg (2200 lb) is reasonable for large adults of A. fragilis, but that 700 kg (1500 lb) is a closer estimate for individuals represented by the average-sized thigh bones he has measured. Using the subadult specimen nicknamed "Big Al", researchers using computer modelling arrived at a best estimate of 1,500 kilograms (3,300 lb) for the individual, but by varying parameters they found a range from approximately 1,400 kilograms (3,100 lb) to approximately 2,000 kilograms (4,400 lb). But more recent studies stated that most Allosaurus(if not all) died young and Allosaurus is surprisingly dense in structure which has boosted the average size for most people to 2,000+ kilograms (4,400+ lb) with the largest reaching almost 3,000 kilograms(6,600 lb). Huge variation not only lies with the weight but the length(especially for the average) is very wide spread. Reasonable estimates range from 8.5 metres to 10 metres on average and some very liberal estimates even take it to 12+ metres on average though its mostly based on Saurophaganax and Epanterias.

Allosaurus was the most successful apex predator in its age and one of the most successful of all time
Discovery and history:
The early years(1800's)
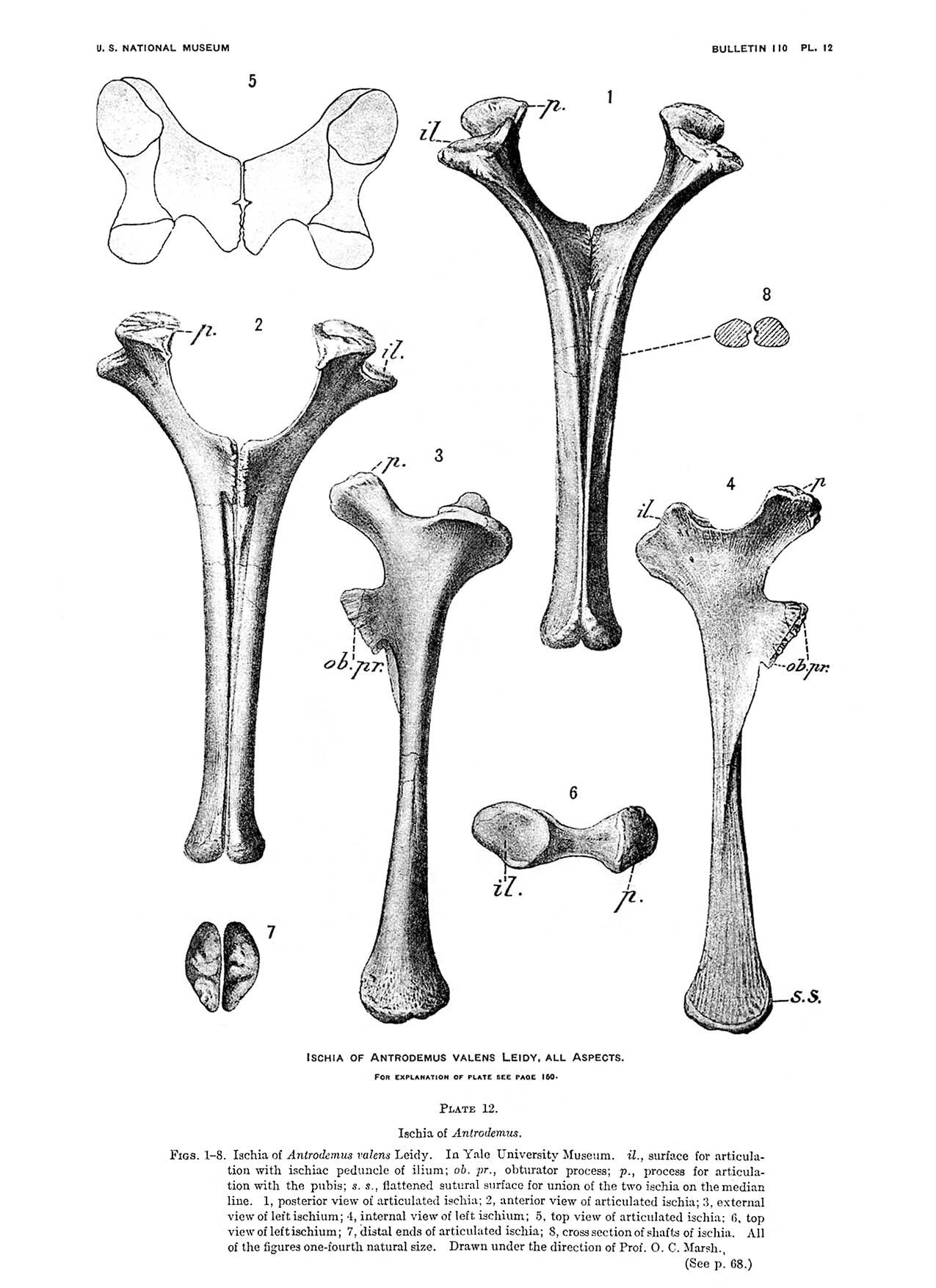
The discovery and early study of Allosaurus is complicated by the multiplicity of names coined during the Bone Wars of the late 19th century. The first described fossil in this history was a bone obtained secondhand by Ferdinand Vandiveer Hayden in 1869. It came from Middle Park, near Granby, Colorado, probably from Morrison Formation rocks. The locals had identified such bones as "petrified horse hoofs". Hayden sent his specimen to Joseph Leidy, who identified it as half of a tail vertebra, and tentatively assigned it to the European dinosaur genus Poekilopleuron as Poicilopleuron valens. He later decided it deserved its own genus, Antrodemus.
The multiplicity of early names complicated later research, with the situation compounded by the terse descriptions provided by Marsh and Cope. Even at the time, authors such as Samuel Wendell Williston suggested that too many names had been coined. For example, Williston pointed out in 1901 that Marsh had never been able to adequately distinguish Allosaurus from Creosaurus. The most influential early attempt to sort out the convoluted situation was produced by Charles W. Gilmore in 1920. He came to the conclusion that the tail vertebra dubbed Antrodemus by Leidy was indistinguishable from those of Allosaurus, and Antrodemus thus should be the preferred name because as the older name it had priority. Antrodemus became the accepted name for this familiar genus for over fifty years, until James Madsen published on the Cleveland-Lloyd specimens and concluded that Allosaurus should be used because Antrodemus was based on material with poor, if any, diagnostic features and locality information (for example, the geological formation that the single bone of Antrodemus came from is unknown). "Antrodemus" has been used informally for convenience when distinguishing between the skull Gilmore restored and the composite skull restored by Madsen.
Cleveland-Lloyd discoveries(1945-1965)
Although sporadic work at what became known as the Cleveland-Lloyd Dinosaur Quarry in Emery County, Utah had taken place as early as 1927, and the fossil site itself described by William J. Stokes in 1945, major operations did not begin there until 1960. Under a cooperative effort involving nearly 40 institutions, thousands of bones were recovered between 1960 and 1965. The quarry is notable for the predominance of Allosaurus remains, the condition of the specimens, and the lack of scientific resolution on how it came to be. The majority of bones belong to the large theropod Allosaurus fragilis (it is estimated that the remains of at least 46 A. fragilis have been found there, out of at minimum 73 dinosaurs), and the fossils found there are disarticulated and well-mixed. Nearly a dozen scientific papers have been written on the taphonomy of the site, suggesting numerous mutually exclusive explanations for how it may have formed. Suggestions have ranged from animals getting stuck in a bog, to becoming trapped in deep mud, to falling victim to drought-induced mortality around a waterhole, to getting trapped in a spring-fed pond or seep. Regardless of the actual cause, the great quantity of well-preserved Allosaurus remains has allowed this genus to be known in detail, making it among the best-known theropods. Skeletal remains from the quarry pertain to individuals of almost all ages and sizes, from less than 1 meter (3.3 ft) to 12 meters (39 ft) long, and the disarticulation is an advantage for describing bones usually found fused.
Recent work and the Big als(1980s–present)
The period since Madsen's monograph has been marked by a great expansion in studies dealing with topics concerning Allosaurus in life (paleobiological and paleoecological topics). Such studies have covered topics including skeletal variation, growth, skull construction, hunting methods, the brain, and the possibility of gregarious living and parental care. Reanalysis of old material (particularly of large 'allosaur' specimens), new discoveries in Portugal, and several very complete new specimens have also contributed to the growing knowledge base.
One of the more significant Allosaurus finds was the 1991 discovery of "Big Al" (MOR 693), a 95% complete, partially articulated specimen that measured about 8 meters (about 26 ft) in length. MOR 693 was excavated near Shell, Wyoming, by a joint Museum of the Rockies and University of Wyoming Geological Museum team. This skeleton was discovered by a Swiss team, led by Kirby Siber. In 1996 the same team discovered a second Allosaurus, "Big Al Two", which is the best preserved skeleton of its kind to date. The completeness, preservation, and scientific importance of this skeleton gave "Big Al" its name; the individual itself was below the average size for Allosaurus fragilis, and was a subadult estimated at only 87% grown. The specimen was described by Breithaupt in 1996. 19 of its bones were broken or showed signs of infection, which may have contributed to "Big Al's" death. Pathologic bones included five ribs, five vertebrae, and four bones of the feet; several damaged bones showed osteomyelitis, a bone infection. A particular problem for the living animal was infection and trauma to the right foot that probably affected movement and may have also predisposed the other foot to injury because of a change in gait.

Allosaurus was the most common large theropod in the vast tract of Western American fossil-bearing rock known as the Morrison Formation, accounting for 70 to 75% of theropod specimens, and as such was at the top trophic level of the Morrison food web. The Morrison Formation is interpreted as a semiarid environment with distinct wet and dry seasons, and flat floodplains. Vegetation varied from river-lining forests of conifers, tree ferns, and ferns (gallery forests), to fern savannah with occasional trees such as the Araucaria-like conifer Brachyphyllum.
The Morrison Formation has been a rich fossil hunting ground. The flora of the period has been revealed by fossils of green algae, fungi, mosses, horsetails, ferns, cycads, ginkgoes, and several families of conifers. Animal fossils discovered include bivalves, snails, ray-finned fishes, frogs, salamanders, turtles, sphenodonts, lizards, terrestrial and aquatic crocodylomorphans, several species of pterosaur, numerous dinosaur species, and early mammals such as docodonts, multituberculates, symmetrodonts, and triconodonts. Dinosaurs known from the Morrison include the theropods Ceratosaurus, Ornitholestes, and Torvosaurus, the sauropods Apatosaurus, Brachiosaurus, Camarasaurus, and Diplodocus, and the ornithischians Camptosaurus, Dryosaurus, and Stegosaurus. It is commonly found at the same sites as Apatosaurus, Camarasaurus, Diplodocus, and Stegosaurus. The Late Jurassic formations of Portugal where Allosaurus is present are interpreted as having been similar to the Morrison but with a stronger marine influence. Many of the dinosaurs of the Morrison Formation are the same genera as those seen in Portuguese rocks (mainly Allosaurus, Ceratosaurus, Torvosaurus, and Apatosaurus), or have a close counterpart (Brachiosaurus and Lusotitan, Camptosaurus and Draconyx).
Allosaurus coexisted with fellow large theropods Ceratosaurus and Torvosaurus in both the United States and Portugal. The three appear to have had different ecological niches, based on anatomy and the location of fossils. Ceratosaurs and torvosaurs may have preferred to be active around waterways, and had lower, thinner bodies that would have given them an advantage in forest and underbrush terrains, whereas allosaurs were more compact, with longer legs, faster but less manoeuvrable, and seem to have preferred dry floodplains. Ceratosaurus, better known than Torvosaurus, differed noticeably from Allosaurus in functional anatomy by having a taller, narrower skull with large, broad teeth. Allosaurus was itself a potential food item to other carnivores, as illustrated by an Allosaurus pubic foot marked by the teeth of another theropod, probably Ceratosaurus or Torvosaurus. The location of the bone in the body (along the bottom margin of the torso and partially shielded by the legs), and the fact that it was among the most massive in the skeleton, indicates that the Allosaurus was being scavenged.

A scene from dinosaur revolution depicting Portugal 155 million years ago.
Allosauridae is one of four families in Carnosauria; the other three are Neovenatoridae, Carcharodontosauridae and Sinraptoridae. Allosauridae has at times been proposed as ancestral to the Tyrannosauridae (which would make it paraphyletic), one recent example being Gregory S. Paul's Predatory Dinosaurs of the World, but this has been rejected, with tyrannosaurids identified as members of a separate branch of theropods, the Coelurosauria. Allosauridae is the smallest of the carnosaur families, with only Saurophaganax and a currently unnamed French allosauroid accepted as possible valid genera besides Allosaurus in the most recent review. Another genus, Epanterias, is a potential valid member, but it and Saurophaganax may turn out to be large examples of Allosaurus. Recent reviews have kept the genus Saurophaganax and included Epanterias with Allosaurus.

Cladogram of the entire carnosauria
Allosaurus fragilis had an average length of 7-9 meters (~30 feet), with the largest definitive Allosaurus specimen (AMNH 680) measuring 9.7 m (32 ft). Several gigantic specimens have been attributed to Allosaurus, but may in fact belong to other genera. The closely related genus Saurophaganax (OMNH 1708) reached 10.9 m (36 ft) in length, and has sometimes been included in the genus Allosaurus as Allosaurus maximus, though recent studies lend support to the idea that it does belong in a seperate genus. Another specimen, once assigned to the genus Epanterias (AMNH 5767), may have measured 12.1 m in length. Epanterias may be a species of Allosaurus or Saurophaganax.
As with dinosaurs in general, weight estimates are debatable, and since 1980 have ranged between 1500 kilograms (3300 lb), 1000 to 4000 kilograms (2200 to 8800 lb), and 1010 kilograms (2230 lb) for modal adult weight (not maximum). John Foster, a specialist on the Morrison Formation, suggests that 1000 kg (2200 lb) is reasonable for large adults of A. fragilis, but that 700 kg (1500 lb) is a closer estimate for individuals represented by the average-sized thigh bones he has measured. Using the subadult specimen nicknamed "Big Al", researchers using computer modelling arrived at a best estimate of 1,500 kilograms (3,300 lb) for the individual, but by varying parameters they found a range from approximately 1,400 kilograms (3,100 lb) to approximately 2,000 kilograms (4,400 lb). But more recent studies stated that most Allosaurus(if not all) died young and Allosaurus is surprisingly dense in structure which has boosted the average size for most people to 2,000+ kilograms (4,400+ lb) with the largest reaching almost 3,000 kilograms(6,600 lb). Huge variation not only lies with the weight but the length(especially for the average) is very wide spread. Reasonable estimates range from 8.5 metres to 10 metres on average and some very liberal estimates even take it to 12+ metres on average though its mostly based on Saurophaganax and Epanterias.
Allosaurus was the most successful apex predator in its age and one of the most successful of all time
Discovery and history:
The early years(1800's)
The discovery and early study of Allosaurus is complicated by the multiplicity of names coined during the Bone Wars of the late 19th century. The first described fossil in this history was a bone obtained secondhand by Ferdinand Vandiveer Hayden in 1869. It came from Middle Park, near Granby, Colorado, probably from Morrison Formation rocks. The locals had identified such bones as "petrified horse hoofs". Hayden sent his specimen to Joseph Leidy, who identified it as half of a tail vertebra, and tentatively assigned it to the European dinosaur genus Poekilopleuron as Poicilopleuron valens. He later decided it deserved its own genus, Antrodemus.
The multiplicity of early names complicated later research, with the situation compounded by the terse descriptions provided by Marsh and Cope. Even at the time, authors such as Samuel Wendell Williston suggested that too many names had been coined. For example, Williston pointed out in 1901 that Marsh had never been able to adequately distinguish Allosaurus from Creosaurus. The most influential early attempt to sort out the convoluted situation was produced by Charles W. Gilmore in 1920. He came to the conclusion that the tail vertebra dubbed Antrodemus by Leidy was indistinguishable from those of Allosaurus, and Antrodemus thus should be the preferred name because as the older name it had priority. Antrodemus became the accepted name for this familiar genus for over fifty years, until James Madsen published on the Cleveland-Lloyd specimens and concluded that Allosaurus should be used because Antrodemus was based on material with poor, if any, diagnostic features and locality information (for example, the geological formation that the single bone of Antrodemus came from is unknown). "Antrodemus" has been used informally for convenience when distinguishing between the skull Gilmore restored and the composite skull restored by Madsen.
Cleveland-Lloyd discoveries(1945-1965)
Although sporadic work at what became known as the Cleveland-Lloyd Dinosaur Quarry in Emery County, Utah had taken place as early as 1927, and the fossil site itself described by William J. Stokes in 1945, major operations did not begin there until 1960. Under a cooperative effort involving nearly 40 institutions, thousands of bones were recovered between 1960 and 1965. The quarry is notable for the predominance of Allosaurus remains, the condition of the specimens, and the lack of scientific resolution on how it came to be. The majority of bones belong to the large theropod Allosaurus fragilis (it is estimated that the remains of at least 46 A. fragilis have been found there, out of at minimum 73 dinosaurs), and the fossils found there are disarticulated and well-mixed. Nearly a dozen scientific papers have been written on the taphonomy of the site, suggesting numerous mutually exclusive explanations for how it may have formed. Suggestions have ranged from animals getting stuck in a bog, to becoming trapped in deep mud, to falling victim to drought-induced mortality around a waterhole, to getting trapped in a spring-fed pond or seep. Regardless of the actual cause, the great quantity of well-preserved Allosaurus remains has allowed this genus to be known in detail, making it among the best-known theropods. Skeletal remains from the quarry pertain to individuals of almost all ages and sizes, from less than 1 meter (3.3 ft) to 12 meters (39 ft) long, and the disarticulation is an advantage for describing bones usually found fused.
Recent work and the Big als(1980s–present)
The period since Madsen's monograph has been marked by a great expansion in studies dealing with topics concerning Allosaurus in life (paleobiological and paleoecological topics). Such studies have covered topics including skeletal variation, growth, skull construction, hunting methods, the brain, and the possibility of gregarious living and parental care. Reanalysis of old material (particularly of large 'allosaur' specimens), new discoveries in Portugal, and several very complete new specimens have also contributed to the growing knowledge base.
One of the more significant Allosaurus finds was the 1991 discovery of "Big Al" (MOR 693), a 95% complete, partially articulated specimen that measured about 8 meters (about 26 ft) in length. MOR 693 was excavated near Shell, Wyoming, by a joint Museum of the Rockies and University of Wyoming Geological Museum team. This skeleton was discovered by a Swiss team, led by Kirby Siber. In 1996 the same team discovered a second Allosaurus, "Big Al Two", which is the best preserved skeleton of its kind to date. The completeness, preservation, and scientific importance of this skeleton gave "Big Al" its name; the individual itself was below the average size for Allosaurus fragilis, and was a subadult estimated at only 87% grown. The specimen was described by Breithaupt in 1996. 19 of its bones were broken or showed signs of infection, which may have contributed to "Big Al's" death. Pathologic bones included five ribs, five vertebrae, and four bones of the feet; several damaged bones showed osteomyelitis, a bone infection. A particular problem for the living animal was infection and trauma to the right foot that probably affected movement and may have also predisposed the other foot to injury because of a change in gait.
The skeleton of big al 2, the most complete allosaurus known to date
Paleoecology:Allosaurus was the most common large theropod in the vast tract of Western American fossil-bearing rock known as the Morrison Formation, accounting for 70 to 75% of theropod specimens, and as such was at the top trophic level of the Morrison food web. The Morrison Formation is interpreted as a semiarid environment with distinct wet and dry seasons, and flat floodplains. Vegetation varied from river-lining forests of conifers, tree ferns, and ferns (gallery forests), to fern savannah with occasional trees such as the Araucaria-like conifer Brachyphyllum.
The Morrison Formation has been a rich fossil hunting ground. The flora of the period has been revealed by fossils of green algae, fungi, mosses, horsetails, ferns, cycads, ginkgoes, and several families of conifers. Animal fossils discovered include bivalves, snails, ray-finned fishes, frogs, salamanders, turtles, sphenodonts, lizards, terrestrial and aquatic crocodylomorphans, several species of pterosaur, numerous dinosaur species, and early mammals such as docodonts, multituberculates, symmetrodonts, and triconodonts. Dinosaurs known from the Morrison include the theropods Ceratosaurus, Ornitholestes, and Torvosaurus, the sauropods Apatosaurus, Brachiosaurus, Camarasaurus, and Diplodocus, and the ornithischians Camptosaurus, Dryosaurus, and Stegosaurus. It is commonly found at the same sites as Apatosaurus, Camarasaurus, Diplodocus, and Stegosaurus. The Late Jurassic formations of Portugal where Allosaurus is present are interpreted as having been similar to the Morrison but with a stronger marine influence. Many of the dinosaurs of the Morrison Formation are the same genera as those seen in Portuguese rocks (mainly Allosaurus, Ceratosaurus, Torvosaurus, and Apatosaurus), or have a close counterpart (Brachiosaurus and Lusotitan, Camptosaurus and Draconyx).
Allosaurus coexisted with fellow large theropods Ceratosaurus and Torvosaurus in both the United States and Portugal. The three appear to have had different ecological niches, based on anatomy and the location of fossils. Ceratosaurs and torvosaurs may have preferred to be active around waterways, and had lower, thinner bodies that would have given them an advantage in forest and underbrush terrains, whereas allosaurs were more compact, with longer legs, faster but less manoeuvrable, and seem to have preferred dry floodplains. Ceratosaurus, better known than Torvosaurus, differed noticeably from Allosaurus in functional anatomy by having a taller, narrower skull with large, broad teeth. Allosaurus was itself a potential food item to other carnivores, as illustrated by an Allosaurus pubic foot marked by the teeth of another theropod, probably Ceratosaurus or Torvosaurus. The location of the bone in the body (along the bottom margin of the torso and partially shielded by the legs), and the fact that it was among the most massive in the skeleton, indicates that the Allosaurus was being scavenged.
A scene from dinosaur revolution depicting Portugal 155 million years ago.
Feeding:
Paleontologists accept Allosaurus as an active predator of large animals. There is dramatic evidence for allosaur attacks on Stegosaurus, including an Allosaurus tail vertebra with a partially healed puncture wound that fits a Stegosaurus tail spike, and a Stegosaurus neck plate with a U-shaped wound that correlates well with an Allosaurus snout. Sauropods seem to be likely candidates as both live prey and as objects of scavenging, based on the presence of scrapings on sauropod bones fitting allosaur teeth well and the presence of shed allosaur teeth with sauropod bones. However, as Gregory Paul noted in 1988, Allosaurus was probably not a predator of fully grown sauropods, unless it hunted in packs, as it had a modestly sized skull and relatively small teeth, and was greatly outweighed by contemporaneous sauropods. Another possibility is that it preferred to hunt juveniles instead of fully grown adults. Research in the 1990s and first decade of the 21st century may have found other solutions to this question. Robert T. Bakker, comparing Allosaurus to Cenozoic sabre-toothed carnivorous mammals, found similar adaptations, such as a reduction of jaw muscles and increase in neck muscles, and the ability to open the jaws extremely wide. Although Allosaurus did not have sabre teeth, Bakker suggested another mode of attack that would have used such neck and jaw adaptations: the short teeth in effect became small serrations on a saw-like cutting edge running the length of the upper jaw, which would have been driven into prey. This type of jaw would permit slashing attacks against much larger prey, with the goal of weakening the victim.
Similar conclusions were drawn by another study using finite element analysis on an Allosaurus skull. According to their biomechanical analysis, the skull was very strong but had a relatively small bite force. By using jaw muscles only, it could produce a bite force of 805 to 2,148 N, less than the values for alligators (13,000 N), lions (4,167 N), and leopards (2,268 N), but the skull could withstand nearly 55,500 N of vertical force against the tooth row. The authors suggested that Allosaurus used its skull like a hatchet against prey, attacking open-mouthed, slashing flesh with its teeth, and tearing it away without splintering bones, unlike Tyrannosaurus, which is thought to have been capable of damaging bones. New estimates debunk this popular theory due to the estimate only measuring jaw muscles and not the skull, teeth and neck muscles. From the 2,148 N to almost 9,000 N(8724 N to be exact).
A biomechanical study published in 2013 by Eric Snively and colleagues found that Allosaurus had an unusually low attachment point on the skull for the longissimus capitis superficialis neck muscle compared to other theropods such as Tyrannosaurus. This would have allowed the animal to make rapid and forceful vertical movements with the skull. The authors found that vertical strikes as proposed by Bakker and Rayfield are consistent with the animal's capabilities. They also found that the animal probably processed carcasses by vertical movements in a similar manner to falcons, such as kestrels: the animal could have gripped prey with the skull and feet, then pulled back and up to remove flesh. This differs from the prey-handling envisioned for theropods with more typical necks, which probably tore flesh with lateral shakes of the skull, similar to crocodilians. In addition, Allosaurus was able to "move its head and neck around relatively rapidly and with considerable control", at the cost of power.

A pair of Allosaurus taking on Camarasaurus, this is an actual predator prey relationship.
Paleontologists accept Allosaurus as an active predator of large animals. There is dramatic evidence for allosaur attacks on Stegosaurus, including an Allosaurus tail vertebra with a partially healed puncture wound that fits a Stegosaurus tail spike, and a Stegosaurus neck plate with a U-shaped wound that correlates well with an Allosaurus snout. Sauropods seem to be likely candidates as both live prey and as objects of scavenging, based on the presence of scrapings on sauropod bones fitting allosaur teeth well and the presence of shed allosaur teeth with sauropod bones. However, as Gregory Paul noted in 1988, Allosaurus was probably not a predator of fully grown sauropods, unless it hunted in packs, as it had a modestly sized skull and relatively small teeth, and was greatly outweighed by contemporaneous sauropods. Another possibility is that it preferred to hunt juveniles instead of fully grown adults. Research in the 1990s and first decade of the 21st century may have found other solutions to this question. Robert T. Bakker, comparing Allosaurus to Cenozoic sabre-toothed carnivorous mammals, found similar adaptations, such as a reduction of jaw muscles and increase in neck muscles, and the ability to open the jaws extremely wide. Although Allosaurus did not have sabre teeth, Bakker suggested another mode of attack that would have used such neck and jaw adaptations: the short teeth in effect became small serrations on a saw-like cutting edge running the length of the upper jaw, which would have been driven into prey. This type of jaw would permit slashing attacks against much larger prey, with the goal of weakening the victim.
Similar conclusions were drawn by another study using finite element analysis on an Allosaurus skull. According to their biomechanical analysis, the skull was very strong but had a relatively small bite force. By using jaw muscles only, it could produce a bite force of 805 to 2,148 N, less than the values for alligators (13,000 N), lions (4,167 N), and leopards (2,268 N), but the skull could withstand nearly 55,500 N of vertical force against the tooth row. The authors suggested that Allosaurus used its skull like a hatchet against prey, attacking open-mouthed, slashing flesh with its teeth, and tearing it away without splintering bones, unlike Tyrannosaurus, which is thought to have been capable of damaging bones. New estimates debunk this popular theory due to the estimate only measuring jaw muscles and not the skull, teeth and neck muscles. From the 2,148 N to almost 9,000 N(8724 N to be exact).
A biomechanical study published in 2013 by Eric Snively and colleagues found that Allosaurus had an unusually low attachment point on the skull for the longissimus capitis superficialis neck muscle compared to other theropods such as Tyrannosaurus. This would have allowed the animal to make rapid and forceful vertical movements with the skull. The authors found that vertical strikes as proposed by Bakker and Rayfield are consistent with the animal's capabilities. They also found that the animal probably processed carcasses by vertical movements in a similar manner to falcons, such as kestrels: the animal could have gripped prey with the skull and feet, then pulled back and up to remove flesh. This differs from the prey-handling envisioned for theropods with more typical necks, which probably tore flesh with lateral shakes of the skull, similar to crocodilians. In addition, Allosaurus was able to "move its head and neck around relatively rapidly and with considerable control", at the cost of power.
A pair of Allosaurus taking on Camarasaurus, this is an actual predator prey relationship.
Classification:
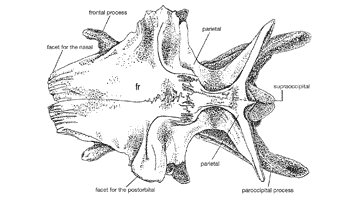
Allosaurus was an allosaurid, a member of a family of large theropods within the larger group Carnosauria. The family name Allosauridae was created for this genus in 1878 by Othniel Charles Marsh, but the term was largely unused until the 1970s in favor of Megalosauridae, another family of large theropods that eventually became a wastebasket taxon. This, along with the use of Antrodemus for Allosaurus during the same period, is a point that needs to be remembered when searching for information on Allosaurus in publications that predate James Madsen's 1976 monograph. Major publications using the name "Megalosauridae" instead of "Allosauridae" include Gilmore, 1920, von Huene, 1926, Romer, 1956 and 1966, Steel, 1970, and Walker, 1964.
Following the publication of Madsen's influential monograph, Allosauridae became the preferred family assignment, but it too was not strongly defined. Semi-technical works used Allosauridae for a variety of large theropods, usually those that were larger and better-known than megalosaurids. Typical theropods that were thought to be related to Allosaurus included Indosaurus, Piatnitzkysaurus, Piveteausaurus, Yangchuanosaurus, Acrocanthosaurus, Chilantaisaurus, Compsosuchus, Stokesosaurus, and Szechuanosaurus. Given modern knowledge of theropod diversity and the advent of cladistic study of evolutionary relationships, none of these theropods is now recognized as an allosaurid, although several, like Acrocanthosaurus and Yangchuanosaurus, are members of closely related families.
Allosaurus was an allosaurid, a member of a family of large theropods within the larger group Carnosauria. The family name Allosauridae was created for this genus in 1878 by Othniel Charles Marsh, but the term was largely unused until the 1970s in favor of Megalosauridae, another family of large theropods that eventually became a wastebasket taxon. This, along with the use of Antrodemus for Allosaurus during the same period, is a point that needs to be remembered when searching for information on Allosaurus in publications that predate James Madsen's 1976 monograph. Major publications using the name "Megalosauridae" instead of "Allosauridae" include Gilmore, 1920, von Huene, 1926, Romer, 1956 and 1966, Steel, 1970, and Walker, 1964.
Following the publication of Madsen's influential monograph, Allosauridae became the preferred family assignment, but it too was not strongly defined. Semi-technical works used Allosauridae for a variety of large theropods, usually those that were larger and better-known than megalosaurids. Typical theropods that were thought to be related to Allosaurus included Indosaurus, Piatnitzkysaurus, Piveteausaurus, Yangchuanosaurus, Acrocanthosaurus, Chilantaisaurus, Compsosuchus, Stokesosaurus, and Szechuanosaurus. Given modern knowledge of theropod diversity and the advent of cladistic study of evolutionary relationships, none of these theropods is now recognized as an allosaurid, although several, like Acrocanthosaurus and Yangchuanosaurus, are members of closely related families.
Allosauridae is one of four families in Carnosauria; the other three are Neovenatoridae, Carcharodontosauridae and Sinraptoridae. Allosauridae has at times been proposed as ancestral to the Tyrannosauridae (which would make it paraphyletic), one recent example being Gregory S. Paul's Predatory Dinosaurs of the World, but this has been rejected, with tyrannosaurids identified as members of a separate branch of theropods, the Coelurosauria. Allosauridae is the smallest of the carnosaur families, with only Saurophaganax and a currently unnamed French allosauroid accepted as possible valid genera besides Allosaurus in the most recent review. Another genus, Epanterias, is a potential valid member, but it and Saurophaganax may turn out to be large examples of Allosaurus. Recent reviews have kept the genus Saurophaganax and included Epanterias with Allosaurus.
Cladogram of the entire carnosauria
Species and taxonomy:
It is unclear how many species of Allosaurus there were. Seven species have been considered potentially valid since 1988 (A. amplexus, A. atrox, A. europaeus, the type species A. fragilis, the as-yet not formally described "A. jimmadseni", A. maximus, and A. tendagurensis), although only a fraction are usually considered valid at any given time. Additionally, there are at least ten dubious or undescribed species that have been assigned to Allosaurus over the years, along with the species belonging to genera now sunk into Allosaurus. In a recent review of basal tetanuran theropods, only A. fragilis (including A. amplexus and A. atrox as synonyms), "A. jimmadseni" (as an unnamed species), and A. tendagurensis were accepted as potentially valid species, with A. europaeus not yet proposed and A. maximus assigned to Saurophaganax.
A. amplexus, A. atrox, A. fragilis, "A. jimmadseni", and A. maximus are all known from remains discovered in the Kimmeridgian–Tithonian Upper Jurassic-age Morrison Formation of the United States, spread across the states of Colorado, Montana, New Mexico, Oklahoma, South Dakota, Utah, and Wyoming. A. fragilis is regarded as the most common, known from the remains of at least sixty individuals. Debate has gone on since the 1980s regarding the possibility that there are two common Morrison Formation species of Allosaurus, with the second known as A. atrox; recent work has followed a "one species" interpretation, with the differences seen in the Morrison Formation material attributed to individual variation. A study of skull elements from the Cleveland-Lloyd site found wide variation between individuals, calling into question previous species-level distinctions based such features as the shape of the lacrimal horns, and the proposed differentiation of "A. jimmadseni" based on the shape of the jugal. A. europaeus was found in the Kimmeridgian-age Porto Novo Member of the Lourinhã Formation, but may be the same as A. fragilis. A. tendagurensis was found in Kimmeridgian-age rocks of Tendaguru, in Mtwara, Tanzania. It may be a more basal tetanuran, a carcharodontosaurid, or simply a dubious theropod. Although obscure, it was a large theropod, possibly around 10 meters long (33 ft) and metric tons 3+ metric tons in weight.
Allosaurus is regarded as a probable synonym of the genera Antrodemus, Creosaurus, Epanterias, and Labrosaurus. Most of the species that are regarded as synonyms of A. fragilis, or that were misassigned to the genus, are obscure and were based on scrappy remains. One exception is Labrosaurus ferox, named in 1884 by Marsh for an oddly formed partial lower jaw, with a prominent gap in the tooth row at the tip of the jaw, and a rear section greatly expanded and turned down. Later researchers suggested that the bone was pathologic, showing an injury to the living animal, and that part of the unusual form of the rear of the bone was due to plaster reconstruction. It is now regarded as an example of A. fragilis. Other remains thought to pertain to Allosaurus have come from across the world, including Australia, Siberia, and Switzerland, but these fossils have been reassessed as belonging to other dinosaurs.
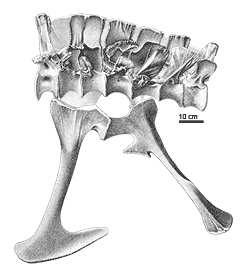
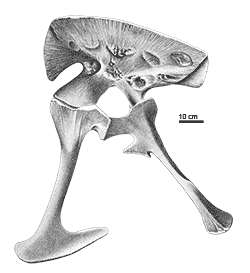
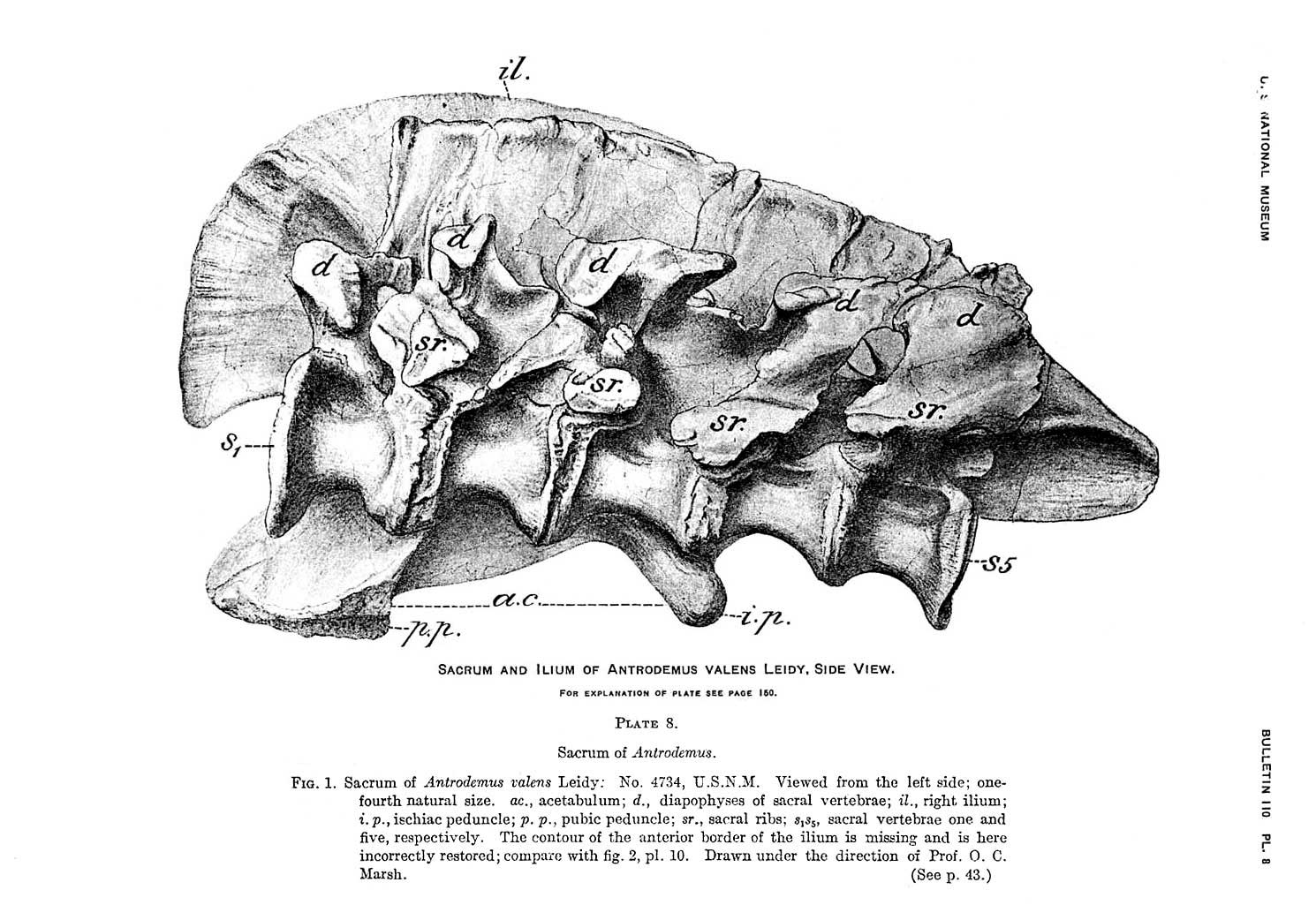
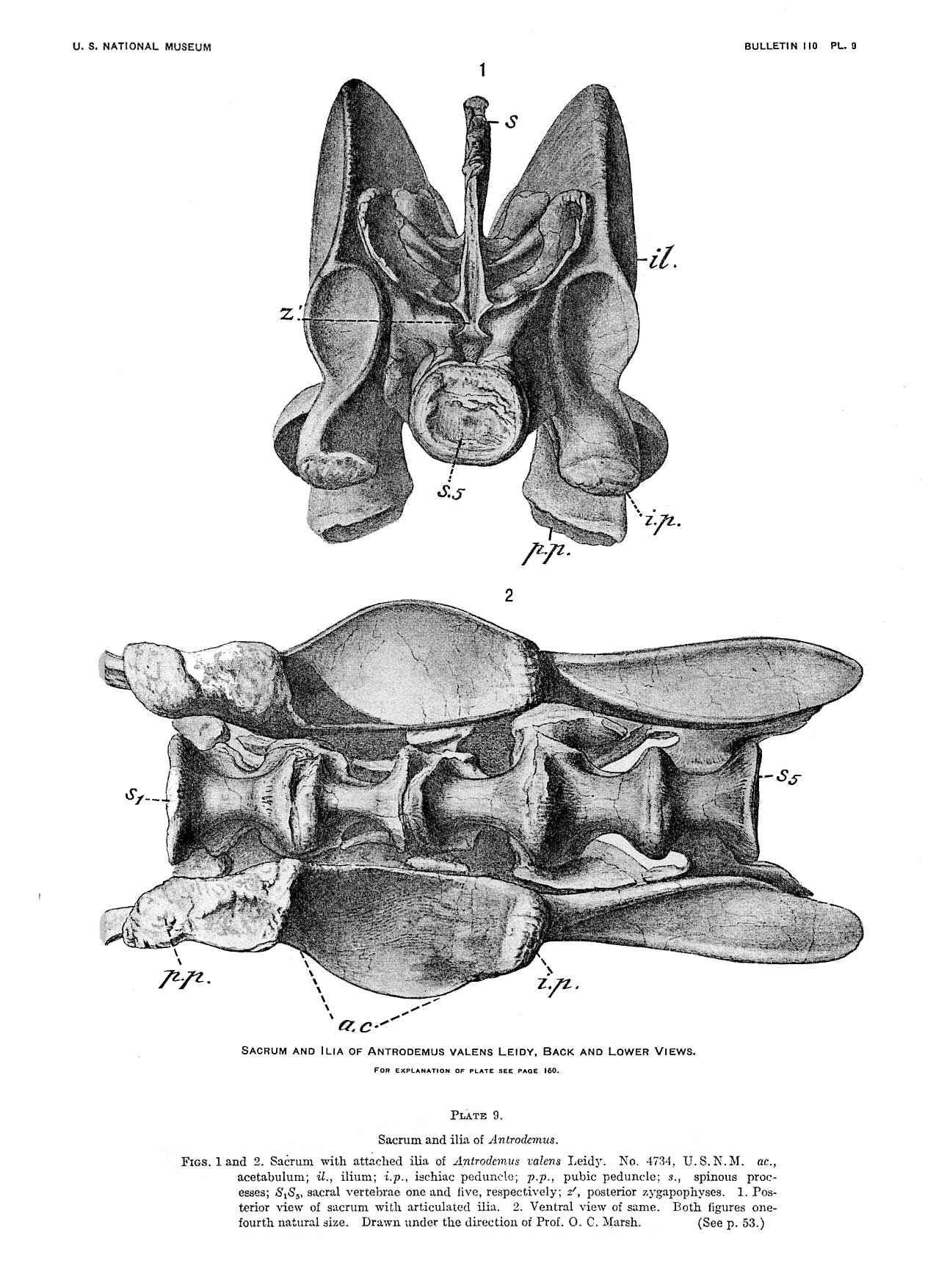
The issue of synonyms is complicated by the type specimen of Allosaurus fragilis (catalog number YPM 1930) being extremely fragmentary, consisting of a few incomplete vertebrae, limb bone fragments, rib fragments, and a tooth. Because of this, several scientists have interpreted the type specimen as potentially dubious, and thus the genus Allosaurus itself or at least the species A. fragilis would be a nomen dubium ("dubious name", based on a specimen too incomplete to compare to other specimens or to classify). To address this situation, Gregory S. Paul and Kenneth Carpenter (2010) submitted a petition to the ICZN to have the name "A. fragilis" officially transferred to the more complete specimen USNM4734 (as a neotype). This request is currently pending review.

It is unclear how many species of Allosaurus there were. Seven species have been considered potentially valid since 1988 (A. amplexus, A. atrox, A. europaeus, the type species A. fragilis, the as-yet not formally described "A. jimmadseni", A. maximus, and A. tendagurensis), although only a fraction are usually considered valid at any given time. Additionally, there are at least ten dubious or undescribed species that have been assigned to Allosaurus over the years, along with the species belonging to genera now sunk into Allosaurus. In a recent review of basal tetanuran theropods, only A. fragilis (including A. amplexus and A. atrox as synonyms), "A. jimmadseni" (as an unnamed species), and A. tendagurensis were accepted as potentially valid species, with A. europaeus not yet proposed and A. maximus assigned to Saurophaganax.
A. amplexus, A. atrox, A. fragilis, "A. jimmadseni", and A. maximus are all known from remains discovered in the Kimmeridgian–Tithonian Upper Jurassic-age Morrison Formation of the United States, spread across the states of Colorado, Montana, New Mexico, Oklahoma, South Dakota, Utah, and Wyoming. A. fragilis is regarded as the most common, known from the remains of at least sixty individuals. Debate has gone on since the 1980s regarding the possibility that there are two common Morrison Formation species of Allosaurus, with the second known as A. atrox; recent work has followed a "one species" interpretation, with the differences seen in the Morrison Formation material attributed to individual variation. A study of skull elements from the Cleveland-Lloyd site found wide variation between individuals, calling into question previous species-level distinctions based such features as the shape of the lacrimal horns, and the proposed differentiation of "A. jimmadseni" based on the shape of the jugal. A. europaeus was found in the Kimmeridgian-age Porto Novo Member of the Lourinhã Formation, but may be the same as A. fragilis. A. tendagurensis was found in Kimmeridgian-age rocks of Tendaguru, in Mtwara, Tanzania. It may be a more basal tetanuran, a carcharodontosaurid, or simply a dubious theropod. Although obscure, it was a large theropod, possibly around 10 meters long (33 ft) and metric tons 3+ metric tons in weight.
Allosaurus is regarded as a probable synonym of the genera Antrodemus, Creosaurus, Epanterias, and Labrosaurus. Most of the species that are regarded as synonyms of A. fragilis, or that were misassigned to the genus, are obscure and were based on scrappy remains. One exception is Labrosaurus ferox, named in 1884 by Marsh for an oddly formed partial lower jaw, with a prominent gap in the tooth row at the tip of the jaw, and a rear section greatly expanded and turned down. Later researchers suggested that the bone was pathologic, showing an injury to the living animal, and that part of the unusual form of the rear of the bone was due to plaster reconstruction. It is now regarded as an example of A. fragilis. Other remains thought to pertain to Allosaurus have come from across the world, including Australia, Siberia, and Switzerland, but these fossils have been reassessed as belonging to other dinosaurs.
The issue of synonyms is complicated by the type specimen of Allosaurus fragilis (catalog number YPM 1930) being extremely fragmentary, consisting of a few incomplete vertebrae, limb bone fragments, rib fragments, and a tooth. Because of this, several scientists have interpreted the type specimen as potentially dubious, and thus the genus Allosaurus itself or at least the species A. fragilis would be a nomen dubium ("dubious name", based on a specimen too incomplete to compare to other specimens or to classify). To address this situation, Gregory S. Paul and Kenneth Carpenter (2010) submitted a petition to the ICZN to have the name "A. fragilis" officially transferred to the more complete specimen USNM4734 (as a neotype). This request is currently pending review.
Scale of some of the supposed allosaurus species
Paleobiology:
Life history: The wealth of Allosaurus fossils, from nearly all ages of individuals, allows scientists to study how the animal grew and how long its lifespan may have been. Remains may reach as far back in the lifespan as eggs—crushed eggs from Colorado have been suggested as those of Allosaurus. Based on histological analysis of limb bones, bone deposition appears to stop at around 22 to 28 years, which is comparable to that of other large theropods like Tyrannosaurus. From the same analysis, its maximum growth appears to have been at age 15, with an estimated growth rate of about 150 kilograms (330 lb) per year.
Medullary bone tissue (endosteally derived, ephemeral, mineralization located inside the medulla of the long bones in gravid female birds) has been reported in at least one Allosaurus specimen, a shin bone from the Cleveland-Lloyd Quarry. Today, this bone tissue is only formed in female birds that are laying eggs, as it is used to supply calcium to shells. Its presence in the Allosaurus individual has been used to establish sex and show it had reached reproductive age. However, other studies have called into question some cases of medullary bone in dinosaurs, including this Allosaurus individual. Data from extant birds suggested that the medullary bone in this Allosaurus individual may have been the result of a bone pathology instead.
The discovery of a juvenile specimen with a nearly complete hindlimb shows that the legs were relatively longer in juveniles, and the lower segments of the leg (shin and foot) were relatively longer than the thigh. These differences suggest that younger Allosaurus were faster and had different hunting strategies than adults, perhaps chasing small prey as juveniles, then becoming ambush hunters of large prey upon adulthood. The thigh bone became thicker and wider during growth, and the cross-section less circular, as muscle attachments shifted, muscles became shorter, and the growth of the leg slowed. These changes imply that juvenile legs has less predictable stresses compared with adults, which would have moved with more regular forward progression. Conversely, the skull bones appear to have generally grown isometrically, increasing in size without changing in proportion.
Social behavior: it has been speculated since the 1970s that Allosaurus preyed on sauropods and other large dinosaurs by hunting in groups. Such a depiction is common in semitechnical and popular dinosaur literature. Robert T. Bakker has extended social behavior to parental care, and has interpreted shed allosaur teeth and chewed bones of large prey animals as evidence that adult allosaurs brought food to lairs for their young to eat until they were grown, and prevented other carnivores from scavenging on the food. However, there is actually little evidence of gregarious behavior in theropods, and social interactions with members of the same species would have included antagonistic encounters, as shown by injuries to gastralia and bite wounds to skulls (the pathologic lower jaw named Labrosaurus ferox is one such possible example). Such head-biting may have been a way to establish dominance in a pack or to settle territorial disputes.
Although Allosaurus may have hunted in packs, it has been argued that Allosaurus and other theropods had largely aggressive interactions instead of cooperative interactions with other members of their own species. The study in question noted that cooperative hunting of prey much larger than an individual predator, as is commonly inferred for theropod dinosaurs, is rare among vertebrates in general, and modern diapsid carnivores (including lizards, crocodiles, and birds) very rarely cooperate to hunt in such a way. Instead, they are typically territorial and will kill and cannibalize intruders of the same species, and will also do the same to smaller individuals that attempt to eat before they do when aggregated at feeding sites. According to this interpretation, the accumulation of remains of multiple Allosaurus individuals at the same site, e.g. in the Cleveland–Lloyd Quarry, are not due to pack hunting, but to the fact that Allosaurus individuals were drawn together to feed on other disabled or dead allosaurs, and were sometimes killed in the process. This could explain the high proportion of juvenile and subadult allosaurs present, as juveniles and subadults are disproportionally killed at modern group feeding sites of animals like crocodiles and Komodo dragons. The same interpretation applies to Bakker's lair sites. There is some evidence for cannibalism in Allosaurus, including Allosaurus shed teeth found among rib fragments, possible tooth marks on a shoulder blade, and cannibalized allosaur skeletons among the bones at Bakker's lair sites.

Life history: The wealth of Allosaurus fossils, from nearly all ages of individuals, allows scientists to study how the animal grew and how long its lifespan may have been. Remains may reach as far back in the lifespan as eggs—crushed eggs from Colorado have been suggested as those of Allosaurus. Based on histological analysis of limb bones, bone deposition appears to stop at around 22 to 28 years, which is comparable to that of other large theropods like Tyrannosaurus. From the same analysis, its maximum growth appears to have been at age 15, with an estimated growth rate of about 150 kilograms (330 lb) per year.
Medullary bone tissue (endosteally derived, ephemeral, mineralization located inside the medulla of the long bones in gravid female birds) has been reported in at least one Allosaurus specimen, a shin bone from the Cleveland-Lloyd Quarry. Today, this bone tissue is only formed in female birds that are laying eggs, as it is used to supply calcium to shells. Its presence in the Allosaurus individual has been used to establish sex and show it had reached reproductive age. However, other studies have called into question some cases of medullary bone in dinosaurs, including this Allosaurus individual. Data from extant birds suggested that the medullary bone in this Allosaurus individual may have been the result of a bone pathology instead.
The discovery of a juvenile specimen with a nearly complete hindlimb shows that the legs were relatively longer in juveniles, and the lower segments of the leg (shin and foot) were relatively longer than the thigh. These differences suggest that younger Allosaurus were faster and had different hunting strategies than adults, perhaps chasing small prey as juveniles, then becoming ambush hunters of large prey upon adulthood. The thigh bone became thicker and wider during growth, and the cross-section less circular, as muscle attachments shifted, muscles became shorter, and the growth of the leg slowed. These changes imply that juvenile legs has less predictable stresses compared with adults, which would have moved with more regular forward progression. Conversely, the skull bones appear to have generally grown isometrically, increasing in size without changing in proportion.
Social behavior: it has been speculated since the 1970s that Allosaurus preyed on sauropods and other large dinosaurs by hunting in groups. Such a depiction is common in semitechnical and popular dinosaur literature. Robert T. Bakker has extended social behavior to parental care, and has interpreted shed allosaur teeth and chewed bones of large prey animals as evidence that adult allosaurs brought food to lairs for their young to eat until they were grown, and prevented other carnivores from scavenging on the food. However, there is actually little evidence of gregarious behavior in theropods, and social interactions with members of the same species would have included antagonistic encounters, as shown by injuries to gastralia and bite wounds to skulls (the pathologic lower jaw named Labrosaurus ferox is one such possible example). Such head-biting may have been a way to establish dominance in a pack or to settle territorial disputes.
Although Allosaurus may have hunted in packs, it has been argued that Allosaurus and other theropods had largely aggressive interactions instead of cooperative interactions with other members of their own species. The study in question noted that cooperative hunting of prey much larger than an individual predator, as is commonly inferred for theropod dinosaurs, is rare among vertebrates in general, and modern diapsid carnivores (including lizards, crocodiles, and birds) very rarely cooperate to hunt in such a way. Instead, they are typically territorial and will kill and cannibalize intruders of the same species, and will also do the same to smaller individuals that attempt to eat before they do when aggregated at feeding sites. According to this interpretation, the accumulation of remains of multiple Allosaurus individuals at the same site, e.g. in the Cleveland–Lloyd Quarry, are not due to pack hunting, but to the fact that Allosaurus individuals were drawn together to feed on other disabled or dead allosaurs, and were sometimes killed in the process. This could explain the high proportion of juvenile and subadult allosaurs present, as juveniles and subadults are disproportionally killed at modern group feeding sites of animals like crocodiles and Komodo dragons. The same interpretation applies to Bakker's lair sites. There is some evidence for cannibalism in Allosaurus, including Allosaurus shed teeth found among rib fragments, possible tooth marks on a shoulder blade, and cannibalized allosaur skeletons among the bones at Bakker's lair sites.
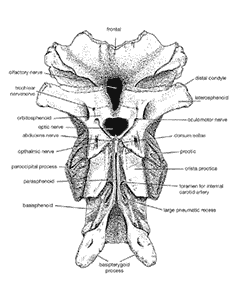
Brain and senses: The cerebrum of Allosaurus, as translated from winding CT examining of an endocast, was more steady with crocodilian brains than those of the other living archosaurs, fledglings. The structure of the vestibular contraption demonstrates that the skull was held almost even, rather than decidedly tipped up or down. The structure of the internal ear was similar to that of a crocodilian, thus Allosaurus most likely could have heard more level frequencies best, and might have had issue with inconspicuous sounds. The olfactory knobs were substantial and appear to have been appropriate for identifying smells, in spite of the fact that the range for assessing scents was generally little.
Paleopathology:
In 2001, Bruce Rothschild and others distributed a study inspecting confirmation for anxiety breaks and tendon separations in theropod dinosaurs and the suggestions for their conduct. Since anxiety cracks are brought on by rehashed trauma as opposed to independent occasions they are less averse to be initiated by the conduct of the creature than different sorts of damage. Anxiety breaks and tendon separations happening in the forelimb have unique behavioral noteworthiness since while wounds to the feet could be created by running or relocation, safe prey things are the most reasonable to assume wellspring of damages to the hand. Allosaurus was one of just two theropods analyzed in the study to display a tendon separation, and in both cases the separation happened on the forelimb. The point when the specialists searched for anxiety cracks, they found that Allosaurus had an altogether more terrific number of anxiety breaks than Albertosaurus, Ornithomimus or Archaeornithomimus. Of the 47 hand bones the specialists concentrated on, 3 were found to hold anxiety cracks. Of the feet, 281 bones were concentrated on and 17 found to have anxiety cracks. The anxiety breaks in the foot bones "were conveyed to the proximal phalanges" and happened over every one of the three weight-bearing toes in "measurably undefined" numbers. Since the more level end of the third metatarsal might have reached the ground first while an allosaur was running it might have borne the most push. In the event that the allosaurs' anxiety cracks were created by harm collecting while strolling or running this bone ought to have encounter a greater number of anxiety breaks than the others. The absence of such a predisposition in the analyzed Allosaurus fossils demonstrates a cause for the anxiety breaks from a source other than running. The creators infer that these cracks happened throughout cooperation with prey, in the same way as an allosaur attempting to hold battling prey with its feet. The richness of anxiety cracks and separation wounds in Allosaurus give confirmation to "exceptionally animated" predation-based instead of searching eating methodologies.
The left scapula and fibula of an Allosaurus fragilis example inventoried as USNM 4734 are both obsessive, both most likely because of mended breaks. The example USNM 8367 saved a few neurotic gastralia which save proof of mended cracks close to their center. A portion of the breaks were inadequately recuperated and "structured pseudoarthroses." An example with a cracked rib was recouped from the Cleveland-Lloyd Quarry. An alternate example had cracked ribs and intertwined vertebrae close to the end of the tail. An evident subadult male Allosaurus fragilis was accounted for to have far reaching pathologies, with what added up to fourteen separate wounds. The example MOR 693 had pathologies on five ribs, the sixth neck vertebra the third eighth and thirteenth back vertebrae, the second tail vertebra and its chevron, the gastralia right scapula, manual phalanx I exited ilium metatarsals III and V, the first phalanx of the third toe and the third phalanx of the second. The ilium had "a substantial gap... brought about by a blow from above".the close end of the first phalanx of the third toe was distressed by an involucrum.
Other pathologies reported in Allosaurus include:
-Willow breaks in two ribs.
-Healed fractures in the humerus and radius.
-Distortion of joint surfaces in the foot possibly due to osteoarthritis or developmental issues.
-Osteopetrosis along the endosteal surface of a tibia.
-Distortions of the joint surfaces of the tail vertebrae possibly due to osetoarthritis or developmental issues.
-Extensive 'neoplastic' ankylosis of caudals," possibly due to physical trauma as well as the fusion of chevrons to centra.
-Coossification of vertebral centra near the end of the tail.
-Amputation of a chevron and foot bone, both possibly a result of bites.
-Extensive exostoses" in the first phalanx of the third toe.
-Lesions similar to those caused by osteomyelitis in two scapulae.
-Bone spurs in a premaxilla, ungual, and two metacarpals.
-Exostosis in a pedal phalanx possibly attributable to an infectious disease.
-A metacarpal with a round depressed fracture.

Paleopathology:
In 2001, Bruce Rothschild and others distributed a study inspecting confirmation for anxiety breaks and tendon separations in theropod dinosaurs and the suggestions for their conduct. Since anxiety cracks are brought on by rehashed trauma as opposed to independent occasions they are less averse to be initiated by the conduct of the creature than different sorts of damage. Anxiety breaks and tendon separations happening in the forelimb have unique behavioral noteworthiness since while wounds to the feet could be created by running or relocation, safe prey things are the most reasonable to assume wellspring of damages to the hand. Allosaurus was one of just two theropods analyzed in the study to display a tendon separation, and in both cases the separation happened on the forelimb. The point when the specialists searched for anxiety cracks, they found that Allosaurus had an altogether more terrific number of anxiety breaks than Albertosaurus, Ornithomimus or Archaeornithomimus. Of the 47 hand bones the specialists concentrated on, 3 were found to hold anxiety cracks. Of the feet, 281 bones were concentrated on and 17 found to have anxiety cracks. The anxiety breaks in the foot bones "were conveyed to the proximal phalanges" and happened over every one of the three weight-bearing toes in "measurably undefined" numbers. Since the more level end of the third metatarsal might have reached the ground first while an allosaur was running it might have borne the most push. In the event that the allosaurs' anxiety cracks were created by harm collecting while strolling or running this bone ought to have encounter a greater number of anxiety breaks than the others. The absence of such a predisposition in the analyzed Allosaurus fossils demonstrates a cause for the anxiety breaks from a source other than running. The creators infer that these cracks happened throughout cooperation with prey, in the same way as an allosaur attempting to hold battling prey with its feet. The richness of anxiety cracks and separation wounds in Allosaurus give confirmation to "exceptionally animated" predation-based instead of searching eating methodologies.
The left scapula and fibula of an Allosaurus fragilis example inventoried as USNM 4734 are both obsessive, both most likely because of mended breaks. The example USNM 8367 saved a few neurotic gastralia which save proof of mended cracks close to their center. A portion of the breaks were inadequately recuperated and "structured pseudoarthroses." An example with a cracked rib was recouped from the Cleveland-Lloyd Quarry. An alternate example had cracked ribs and intertwined vertebrae close to the end of the tail. An evident subadult male Allosaurus fragilis was accounted for to have far reaching pathologies, with what added up to fourteen separate wounds. The example MOR 693 had pathologies on five ribs, the sixth neck vertebra the third eighth and thirteenth back vertebrae, the second tail vertebra and its chevron, the gastralia right scapula, manual phalanx I exited ilium metatarsals III and V, the first phalanx of the third toe and the third phalanx of the second. The ilium had "a substantial gap... brought about by a blow from above".the close end of the first phalanx of the third toe was distressed by an involucrum.
Other pathologies reported in Allosaurus include:
-Willow breaks in two ribs.
-Healed fractures in the humerus and radius.
-Distortion of joint surfaces in the foot possibly due to osteoarthritis or developmental issues.
-Osteopetrosis along the endosteal surface of a tibia.
-Distortions of the joint surfaces of the tail vertebrae possibly due to osetoarthritis or developmental issues.
-Extensive 'neoplastic' ankylosis of caudals," possibly due to physical trauma as well as the fusion of chevrons to centra.
-Coossification of vertebral centra near the end of the tail.
-Amputation of a chevron and foot bone, both possibly a result of bites.
-Extensive exostoses" in the first phalanx of the third toe.
-Lesions similar to those caused by osteomyelitis in two scapulae.
-Bone spurs in a premaxilla, ungual, and two metacarpals.
-Exostosis in a pedal phalanx possibly attributable to an infectious disease.
-A metacarpal with a round depressed fracture.
The broken jaw from dinosaur revolution is pictured above. It shows how resiliant these things were. Surviving for years with a broken jaw.
Papers:
1.Variation in a Population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA
Abstract: "Ontogenetic and individual variation in a population of the large theropod Allosaurus cf. A. fragilis was examined using the Björk superimposition method on the premaxillae, maxillae, lachrymal, postorbital, and jugal. This method allows utilization of incomplete specimens without having to estimate (fabricate) missing data. In addition, it visually shows regions of ontogenetic change and variation. Comparison of these results with other morphometric studies of the same specimens show that most allometric trends are not supported. Instead, ontogenetic change is mostly isometric. In addition, the wide range of individual variation in all the skull elements analyzed makes some autapomorphic characters used to define other theropod taxa suspect."
link: www.bioone.org/doi/abs/10.2517/1342-8144-14.4.250
2. First definitive association between embryonic Allosaurus bones and prismatoolithus eggs in the Morrison Formation (Upper Jurassic, Wyoming, USA)
Abstract: "Despite more than a century of collecting, resulting in one of the best-studied vertebrate fossil records anywhere in the world, the Upper Jurassic Morrison Formation has produced surprisingly few examples of dinosaur eggs associated with embryonic remains. Even more puzzling, none of these seem to pertain to the theropod Allosaurus, one of the most common and best-understood dinosaur taxa in the formation. Here we report on a dinosaur nest site that has produced both abundant prismatoolithid eggshell and embryonic (or perinatal) bones of Allosaurus from Fox Mesa, Wyoming. This represents the first such discovery for any theropod in the Jurassic of North America. The nest is heavily weathered but contains a few ellipsoid eggshell clusters that suggest an egg size of about 8 x 6.5 cm. Study of the eggshell morphology and microstructure confirms that a single egg type is present throughout, which is indistinguishable from Prismatoolithus coloradensis. All of the identifiable embryonic materials pertain to theropods, and two premaxillae specimens show the five alveoli diagnostic for Allosaurus among Morrison theropods. This confirms the theropod origin of Prismatoolithus eggs and implicates Allosaurus as the specific Morrison parent taxon. As a result, it is now possible to assign several previous discoveries of dinosaur eggs and potential nests to Allosaurus, including the isolated egg from the Cleveland-Lloyd Quarry. This discovery also calls into question prior assignments of Prismatoolithus eggs to ornithopods, and suggests that more detailed study of such sites is warranted. Prismatoolithus eggshells are also associated with the Upper Jurassic theropod Lourinhanosaurus from Portugal, along with larger embryos that exhibit four premaxillary alveoli."
Link: docentes.fct.unl.pt/omateus/publications/first-definitive-association-between-embryonic-allosaurus-bones-and-prismatooli
3. The Discovery of a Nearly Complete Allosaurus from the Jurassic Morrison Formation, Eastern Bighorn Basin, Wyoming
Abstract: "The carnivorous dinosaur Allosaurus has been known for over 100 years, but in 1991 a partially articulated, sub-adult (informally known as "Big Al") of this genus was discovered that is one of the most complete specimens ever found. The well-preserved skeleton was discovered near Shell, Wyoming in the Upper Jurassic Morrison Formation. This area of northern Wyoming has been known to contain dinosaur remains since crews from the American Museum of Natural History in New York collected at the Howe Quarry in the 1930s. Just north of this quarry, "Big Al" was found by crews from Siber & Siber, Inc. on public lands. The specimen is being studied primarily by researchers from Montana State University and the University of Wyoming. Research on this specimen and the area in which it was found is providing exciting new information on Jurassic carnosaurs, as well as the environment in which they lived."
Link: archives.datapages.com/data/wga/data/060/060001/309_wga0600309.htm
1.Variation in a Population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA
Abstract: "Ontogenetic and individual variation in a population of the large theropod Allosaurus cf. A. fragilis was examined using the Björk superimposition method on the premaxillae, maxillae, lachrymal, postorbital, and jugal. This method allows utilization of incomplete specimens without having to estimate (fabricate) missing data. In addition, it visually shows regions of ontogenetic change and variation. Comparison of these results with other morphometric studies of the same specimens show that most allometric trends are not supported. Instead, ontogenetic change is mostly isometric. In addition, the wide range of individual variation in all the skull elements analyzed makes some autapomorphic characters used to define other theropod taxa suspect."
link: www.bioone.org/doi/abs/10.2517/1342-8144-14.4.250
2. First definitive association between embryonic Allosaurus bones and prismatoolithus eggs in the Morrison Formation (Upper Jurassic, Wyoming, USA)
Abstract: "Despite more than a century of collecting, resulting in one of the best-studied vertebrate fossil records anywhere in the world, the Upper Jurassic Morrison Formation has produced surprisingly few examples of dinosaur eggs associated with embryonic remains. Even more puzzling, none of these seem to pertain to the theropod Allosaurus, one of the most common and best-understood dinosaur taxa in the formation. Here we report on a dinosaur nest site that has produced both abundant prismatoolithid eggshell and embryonic (or perinatal) bones of Allosaurus from Fox Mesa, Wyoming. This represents the first such discovery for any theropod in the Jurassic of North America. The nest is heavily weathered but contains a few ellipsoid eggshell clusters that suggest an egg size of about 8 x 6.5 cm. Study of the eggshell morphology and microstructure confirms that a single egg type is present throughout, which is indistinguishable from Prismatoolithus coloradensis. All of the identifiable embryonic materials pertain to theropods, and two premaxillae specimens show the five alveoli diagnostic for Allosaurus among Morrison theropods. This confirms the theropod origin of Prismatoolithus eggs and implicates Allosaurus as the specific Morrison parent taxon. As a result, it is now possible to assign several previous discoveries of dinosaur eggs and potential nests to Allosaurus, including the isolated egg from the Cleveland-Lloyd Quarry. This discovery also calls into question prior assignments of Prismatoolithus eggs to ornithopods, and suggests that more detailed study of such sites is warranted. Prismatoolithus eggshells are also associated with the Upper Jurassic theropod Lourinhanosaurus from Portugal, along with larger embryos that exhibit four premaxillary alveoli."
Link: docentes.fct.unl.pt/omateus/publications/first-definitive-association-between-embryonic-allosaurus-bones-and-prismatooli
3. The Discovery of a Nearly Complete Allosaurus from the Jurassic Morrison Formation, Eastern Bighorn Basin, Wyoming
Abstract: "The carnivorous dinosaur Allosaurus has been known for over 100 years, but in 1991 a partially articulated, sub-adult (informally known as "Big Al") of this genus was discovered that is one of the most complete specimens ever found. The well-preserved skeleton was discovered near Shell, Wyoming in the Upper Jurassic Morrison Formation. This area of northern Wyoming has been known to contain dinosaur remains since crews from the American Museum of Natural History in New York collected at the Howe Quarry in the 1930s. Just north of this quarry, "Big Al" was found by crews from Siber & Siber, Inc. on public lands. The specimen is being studied primarily by researchers from Montana State University and the University of Wyoming. Research on this specimen and the area in which it was found is providing exciting new information on Jurassic carnosaurs, as well as the environment in which they lived."
Link: archives.datapages.com/data/wga/data/060/060001/309_wga0600309.htm




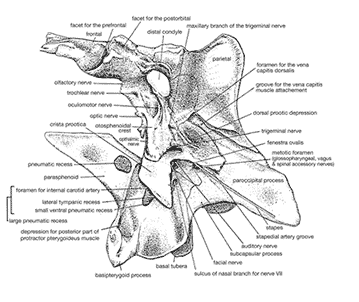
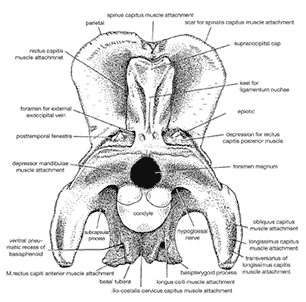
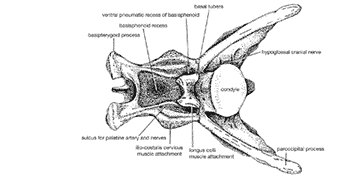
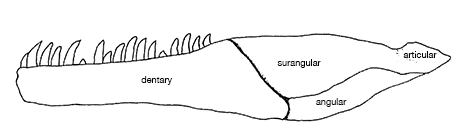






 <- imma rek u btichs
<- imma rek u btichs